crecimiento bacteriano
domingo, 26 de octubre de 2014
Dilucion Bacteriana
Técnica de Recuento por Dilución
Mediante el uso de esta técnica de simple ejecución y fácil interpretación, se pueden evaluar diferentes grupos bacterianos según el medio de cultivo que contenga el frasco.
Es aplicable para evaluar bacterias heterotróficas, aerobias totales (BAT), bacterias reductoras de sulfato (BRS), anaerobias heterotróficas totales (BAnT), etc.
Esta técnica está recomendada por el Instituto Americano del Petróleo (API) para evaluar calidad biológica en aguas de inyección en recuperación secundaria y también es ampliamente usada en evaluación de problemas microbiológicos en aguas de refrigeración.
Procedimiento.
La técnica de dilución por extinción consiste en inocular una serie de frascos tipo antibiótico que contienen 9 ml del medio de cultivo apropiado según el tipo de bacteria que se quiera evaluar.
Se obtiene 1 ml de la muestra usando una jeringa descartable, estéril, de 1 o 2 ml de capacidad.
Se inocula el primer frasco, sin retirar la aguja se agita el mismo, se invierte y se retira 1 ml que se inocula en el 2° frasco.
Con una nueva jeringa, se retira 1 ml del frasco N° 2, previamente agitado, y se inocula el frasco N° 3 y así sucesivamente, utilizando una nueva jeringa para cada frasco.
 Si no existe información previa sobre cantidad de bacterias/mL de la muestra suelen sembrarse 6 frascos.
Si no existe información previa sobre cantidad de bacterias/mL de la muestra suelen sembrarse 6 frascos.
La manifestación de crecimiento bacteriano se observa por la turbidez del medio de cultivo que se compara por transparencia con un frasco sin inocular en el caso de bacterias aerobias totales y bacterias anaerobias totales; en el caso de bacterias reductoras de sulfatos se debe observar un ennegrecimiento del frasco.
Una vez inoculados los frascos y rotulados convenientemente, se incuban durante 7 días (BAT, BAT-RF, MFT) o 14 días (BPA) en estufa de cultivo a ±5ºC respecto de la temperatura del agua muestreada,
efectuando lecturas diarias para observar aquellos frascos que manifiestan desarrollo positivo. Para el caso de los medios BRS y BRH, el plazo de lectura es de 28 días máximo, realizando también lecturas periódicas..
Cálculos de densidad bacteriana.
Se tienen en cuenta los frascos que manifiesten alguna modificación como consecuencia del crecimiento bacteriano; esta modificación puede ser turbidez, cambio de color, ennegrecimiento, etc.
Por ejemplo: si se usó el medio BAT-RF y se observa turbidez y/o cambio de color hasta el frasco N° 4 el resultado sería 1000-10000 bact/mL o exponencialmente 103-104 bact/mL.. Habitualmente se informa 104 bact/mL.
Problemas comunes en la interpretación del crecimiento bacteriano cuando se utiliza la técnica de dilución por extinción.
Hay una variedad de acontecimientos que interfieren con la interpretación de resultados obtenidos utilizando las diluciones seriadas. Algunos de los mas comunes son:
· Todos los frascos muestran crecimiento.
Si esto sucede, no es posible estimar la población, por ejemplo si en una serie de 6 frascos inoculados todos se manifiestan positivos se debe informar:106 bact/mL. Para el próximo análisis de esa muestra habría que considerar el empleo de 8 o 9 frascos.
· Un frasco no denota crecimiento y el resto sí.
Algunas veces uno de los frascos permanece sin cambios mientras que los ubicados anterior y posteriormente en la serie inoculada muestran positividad. Es decir, hay un “salto” en los frascos positivos. Este “salto” debe ser anotado y se informará la concentración bacteriana correspondiente al frasco de mayor numeración que resultó positivo. Por ejemplo: El crecimiento positivo se observa en los frascos 1, 2 y 4 mientras los frascos 3, 5 y 6 permanecen sin modificaciones, todo después de incubar a la temperatura y tiempo incubado. Hay varias explicaciones posibles para esto, incluyendo una contaminación accidental del frasco 4. Por lo tanto, nunca se puede saber que ocurrió realmente y hay que proceder de la forma más simple ante una situación dudosa.
En el ejemplo anterior, si se confía razonablemente en que los frascos fueron rotulados apropiadamente con su número de orden, el informe será 104 bact/mL anotando el frasco que “saltó”.
Características de los cultivos puros
Las colonias deben ser iguales en forma, tamaño y color.
Que no existan formas inhibidas.
Al microscopio deben tener un aspecto común.
Tiene que tener iguales propiedades tintoriales
Deben ser bioquímicamente y fisiológicamente iguales..
Una regla importante es no coger nunca de la primera colonia.
Técnica de la siembra
La técnica para el aislamiento comienza preparando un inóculo de siembra, que es la deposición de una pequeña porción de la muestra en el medio o en los medios de cultivo adecuado, en función de las especies microbianas que se espera encontrar. Por eso existen distintos métodos o tipos de siembra, en función de la muestra de partida, del medio en el que se siembra y de la finalidad del estudio. Describiremos la siembra para inoculación y la siembra para aislamiento.
Siembra para inoculación: la inoculación consiste en tomar una pequeña porción de la muestra y diluirla para dispersarla y que crezca con más facilidad en el medio de cultivo. Se puede inocular en medio líquido o en medio sólido.
Siembra para aislamiento: la finalidad de este tipo de siembras es la de obtener cultivos puros. Se siembra en placa de Petri que contienen medios de cultivo sólidos.
Existen varios tipos de siembras para aislamiento:Siembra en estría o zig-zag, siembra por estría múltiple, siembra de los cuatro cuadrantes, siembra de los tres giros.
Métodos para obtener los cultivos puros
Método de siembra para aislamiento por agotamiento o en estría
Materiales
Placa petri con medio de cultivo
Muestra (sólida o líquida)
Asa de siembra de platino
Con el asa estéril tomamos la muestra y hacemos extensiones en la placa. Volvemos a esterilizar el asa de siembra y extendemos parte de la extensión anterior en otra dirección. Volvemos a esterilizar el asa de siembra y volvemos a extender parte de la segunda extensión en otra dirección. Esterilizamos y repetimos la operación. Incubamos a 37 º durante 24 horas y, en la primera zona, habrá un amasijo de células. En la segunda, parte de la primera zona estará mejor extendida, y en la tercera y en la 4ª mejor aún. Si tomamos una colonia y la ponemos sobre una placa petri, tendremos un cultivo puro.
Estría única: depositamos la muestra lo más alejado de nosotros y realizamos la siembra en forma de zig-zag. De esta forma vamos descargando la muestra del asa. Es importante no volver a repasar el trozo de medio de cultivo que ya hemos sembrado. Para conseguir que al final de la estría tengamos colonias aisladas es importante que al principio hagamos la estría más junta y que la cantidad de muestra tomada en el asa sea la adecuada.
Estría múltiple: en esta técnica de aislamiento por agotamiento se realizan varias estrías. Esto se puede llevar a cabo flameando el asa de paso en paso, es decir, de estría en estría, o sin flamear el asa. Además, dependiendo de la forma en la que hagamos las estrías, existen varios tipos de estriados:
Estriado sobre cuatro cuadrantes: Se utiliza cuando queremos conservar algún microorganismo que nos interesa. Consiste en sembrar uno a uno los cuadrantes, sin flamear el asa entre estría y estría, de tal forma que en el cuarto cuadrante, al ir agotando la muestra del asa, tendremos los microorganismos aislados.
Esterilizando asa entre siembra y siembra: consiste en realizar un primer estriado en uno de los extremos de la placa, esterilizar el asa, girar un poco la placa y realizar otro estriado introduciendo el asa en el primer estriado para ir arrastrando la muestra y volver a repetir esta operación con un tercer estriado:
Sin esterilizar el asa entre siembra y siembra: Se lleva a cabo igual que en el caso anterior salvo en que en esta técnica no se flamea el asa entre la realización de los estriados y por lo tanto no es necesario arrastrar la muestra de las primeras siembras.

Método de siembra para aislamiento por diluciones sucesivas
Consiste en diluir una muestra hasta conseguir muy pocas células que, tras inocularse en un medio de cultivo, generen colonias aisladas.
Metodología: ahora tomamos 0.1 ml de cada tubo y lo depositamos en una placa petri con medio de cultivo. Con un asa de vidrio se extiende esa cantidad sobre la placa, e incubams a 37 ºC durante 24 horas.
En la primera dilución habrá un amasijo de colonias. En la siguiente habrá diez veces menos número de colonias, y así sucesivamente hasta conseguir colonias aisladas. Este método nos permite calcular el número de microorganismos viables que existen en la muestra que estamos analizando: N=C · 1/D · 1/i
donde:
N es el número de microorganismos presentes en la muestra
C es el número de colonias contadas UFC (unidades formadoras de colonias)
D es la dilución
I es el inóculo
Debe haber entre 30 y 300 colonias. Si hay menos, no es significativo (puede tratarse de una contaminación) y, si hay más, no están aisladas.
Técnica del cultivo por enriquecimiento
Consiste en ajustar las condiciones de cultivo para seleccionar el microorganismo que nos interesa aislar. Ejemplo: nos interesa aislar bacterias fotoautótrofas. Tomamos un matraz con agua y sales y lo incubamos a la luz, de manera que crezcan las bacterias fotoautótrofas. Ejemplo: queremos aislar bacterias quimiolitótrofas del azufre; ponemos azufre, CO2 como fuente de carbono (disuelto en agua) e incubamos en la oscuridad para eliminar fotoautótrofos. Para incubar enterobactrias como enterobacter hay que incubar a 37º, mientras que para incubar otra enterobacteria, E. Coli, hay que incubar a 45º.
Conservación de cultivos puros
Almacenamiento a temperaturas bajas. Ponemos las bacterias con glicerol al 20 - 50% a -70º y podemos mantenerlas así durante años y décadas.
Liofilización: es un método más eficaz. En un vial de liofilización ponemos leche desnatada y las bacterias, lo congelamos, lo llevamos al liofilizador y lo sometemos al proceso de liofilización. La liofilización es una evaporación por condensación. Se basa en el punto triple del agua; podemos pasar de sólido a gas. El agua se evapora y queda un polvillo formado por la leche y las bacterias. Cerramos el bote y podemos almacenar el cultivo durante décadas.
Colecciones de cultivo: en todos los países hay organizaciones que preparan o almacenan colecciones de cultivos puros o tipo. En España existe la C.E.C.T (Colección española de cultivos tipo), que está en Valencia. La más grande del mundo es la americana, A.T.C.C. (American type cultive collection).
Mantenimiento
Debemos transferir periódicamente, ya que los tubos de agar se secan, se evapora el agua.
Se debe cubrir con parafina, con lo cual la transferencia se puede hacer más tarde, disminuye el metabolismo y tardan en envejecer.
Se disminuye la temperatura, congelamos las células. Para proteger las células y que no se formen cristales que las dañen se usa el glicerol
La liofilización es la sublimación a alto vacío de las muestras congeladas con rapidez.
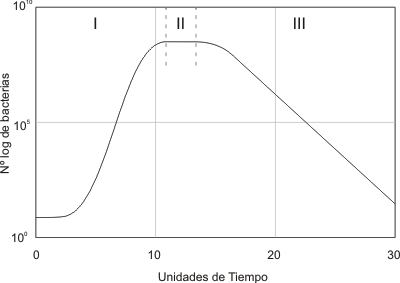
Curvas de Crecimiento Microbiano
El concepto asociado al Crecimiento Microbiano, corresponde al aumento poblacional de una especie microbiana en un medio de cultivo provisto de todas las necesidades del microorganismo (Cantidad de Nutrientes, Temperatura, Grado de Humedad, Gases y pH).
Las curvas de Crecimiento Microbiano constan de 4 etapas bien definidas, aunque el tiempo de duración de cada una de estas etapas, puede variar según el tipo de microorganismo, la familia a la cual este pertenece, entre otras características.
Fase de Latencia: Corresponde a un período de transición para los microorganismos cuando son transferidos a una nueva condición. En esta fase no hay incremento en el número de células, aunque sí una gran actividad en el metabolismo.
Fase de Crecimiento Exponencial: Período en que el crecimiento del microorganismo ocurre de forma exponencial, es decir, cada vez que pasa un determinado tiempo la población se duplica.
Fase Estacionaria: Período en que ocurren las limitaciones del crecimiento, ya sea por agotamiento de algún nutriente esencial, por acumulación de productos tóxicos o por una combinación de las causas anteriores.
Fase de Muerte: Luego que culmine la fase estacionaria, comienza una progresiva disminución en el número de celulas viables, cuando esto ocurre se dice que la población ha entrado en fase de muerte.
TIPOS DE CURVAS DE CRECIMIENTO MICROBIANO


Pruebas bioquímicas
En el laboratorio disponemos de numerosas técnicas para la caracterización bioquímica de una especie
Kligler Hierro Agar Medio de cultivo frecuentemente usado en microbiología de alimentos para la diferenciación de enterobacterias, en base a la fermentación de glucosa y lactosa, y a la producción de ácido sulfhídrico. Fórmula (en gramos por litro) Instrucciones Peptona de carne 13.0 Cloruro de sodio 5.0 Suspender 54.8 g del polvo por litro de Lactosa 10.0 agua destilada. Mezclar bien y calentar Tripteina 10.0 con agitación frecuente, hervir 1 o 2 minutos hasta disolución total. Llenar Glucosa 1.0 hasta la tercera parte de los tubos de Citrato de hierro y amonio 0.5 ensayo. Esterilizar a 121 C por 15 Tiosulfato de sodio 0.3 minutos. Enfriar en pico de flauta profundo. Rojo de fenol 0.025 Agar 15.0 pH final: 7.3 0.2
TSI Agar Medio universalmente empleado para la diferenciación de enterobacterias, en base a la fermentación de glucosa, lactosa, sacarosa y a la producción de ácido sulfhídrico. Fórmula (en gramos por litro) Instrucciones Extracto de carne 3.0 Pluripeptona 20.0 Cloruro de sodio 5.0 Suspender 62,5 g del polvo por litro Lactosa 10.0 de agua destilada. Mezclar bien y calentar con agitación frecuente, Sacarosa 10.0 hervir 1 o 2 minutos hasta disolución Glucosa 1.0 total. Llenar hasta la tercera parte de los tubos de ensayo. Esterilizar a Sulfato de hierro y amonio 0.2 121 C por 15 minutos. Enfriar en pico Tiosulfato de sodio 0.2 de flauta profundo. Rojo de fenol 0.025 Agar 13.0 pH final: 7.3 0.2
Lisina Hierro Agar Medio de cultivo utilizado para diferenciar microorganismos, especialmente Salmonella spp., basado en la decarboxilación / desaminación de la lisina y en la producción de ácido sulfhídrico. Fórmula (en gramos por litro) Instrucciones Peptona de gelatina 5.0 Suspender 35 g del medio Extracto de levadura 3.0 deshidratado en un litro de agua Glucosa 1.0 destilada. Dejar embeber unos 15 minutos. Calentar cuidadosamente, Lisina 10.0 agitando con frecuencia y hervir Citrato de hierro y amonio 0.5 durante un minuto hasta la disolución completa. Distribuir y esterilizar a Tiosulfato de sodio 0.04 121 C durante 15 minutos. Enfriar en Púrpura de bromocresol 0.02 pico de flauta dejando un fondo vertical apto para la punción. Agar 15.0 pH final: 6.7 0.2
MIO Medio Medio usado para la identificación de Enterobacteriaceae en base a su movilidad, actividad de ornitina decarboxilasa y producción de indol. Fórmula (en gramos por litro) Instrucciones Dextrosa 1.0 Extracto de levadura 3.0 Suspender 31 g del polvo en un Peptona 10.0 litro de agua destilada. Calentar Tripteína 10.0 a ebullición hasta completa Clorhidrato de L-ornitina 5.0 disolución. Distribuir en tubos y esterilizar 15 minutos a 121 C. Agar 2.0 Púrpura de bromocresol 0.02 pH final: 6.5 0.2
SIM Medio Es un medio semisólido destinado a verificar la movilidad, producción de indol y de sulfuro de hidrógeno en un mismo tubo. Es útil para diferenciar miembros de la familia Enterobacteriaceae. Fórmula (en gramos por litro) Instrucciones Tripteína 20.0 Suspender 30 g del polvo por litro de agua Peptona 6.1 destilada. Mezclar hasta disolver; calentar agitando y hervir durante un minuto. Distribuir Sulfato de hierro y amonio 0.2 unos 4 ml en tubos de hemólisis y esterilizar Tiosulfato de sodio 0.2 en autoclave a 121 C durante 15 minutos. Agar 3.5 Solidificar en posición vertical. pH final: 7.3 0.2
Urea Medio utilizado para la identificación de microorganismos, en base a la actividad ureásica. Es particularmente útil para diferenciar Proteus spp. de otros miembros de la familia Enterobacteriaceae. Fórmula (en gramos por litro) Instrucciones Urea 20.0 Suspender 3, 87 g del medio Fosfato monopotásico 9.1 deshidratado por cada 100 ml de agua destilada. Disolver sin calentar Fosfato disódico 9.5 y esterilizar por filtración. Distribuir Extracto de levadura 0.1 en tubos pequeños estériles, entre 0,5 y 2 ml. Rojo fenol 0.01 pH final: 6.8 ± 0.2
Simmons Citrato Agar Medio utilizado para la diferenciación de enterobacterias, en base a la capacidad de usar citrato como única fuente de carbono y energía. Fórmula (en gramos por litro) Instrucciones Citrato de sodio 2.0 Cloruro de sodio 5.0 Suspender 24,2 g del medio deshidratado por litro de agua destilada. Fosfato dipotásico 1.0 Dejar reposar 5 minutos y mezclar Fosfato monoamónico 1.0 calentando a ebullición durante 1 o 2 Sulfato de magnesio 0.2 minutos. Distribuir en tubos y esterilizar en autoclave a 121 C durante 15 Azul de bromotimol 0.08 minutos. Enfriar en posición inclinada. Agar 15.0 pH final: 6.9 0.2
sábado, 18 de octubre de 2014
lunes, 13 de octubre de 2014
domingo, 12 de octubre de 2014


TÉCNICAS Y MÉTODOS DE TINCION
La tinción es un método sencillo para incrementar el contraste entre la célula y
su entorno y por lo tanto contribuye a mejorar la imagen observada. Las técnicas de
tinción con diversos colorantes facilitan la observación al aumentar notablemente el
contraste.
En Microbiología todas las tinciones se realizan a partir de suspensiones de
microorganismos extendidas en un portaobjetos (frotis), secadas y fijadas. La fijación,
procedimiento que permite preservar estructuras celulares, se puede llevar a cabo condiferentes tratamientos: fijación por calor o fijación química. La fijación por calor es la
más utilizada para la observación de bacterias. Este procedimiento consiste en pasar el
portaobjetos, con la suspensión bacteriana extendida y seca, a través de una llama de
un mechero. La fijación por calor preserva la morfología externa de los
microorganismos pero no las estructuras internas. La fijación química con agentes
como etanol, folmaldehido y ácido acético entre otros muchos, se utiliza para
preservar las estructuras celulares.
Se pueden utilizar dos tipos de procedimientos, la tinción positiva es la más
frecuente, en ella un colorante se une a ciertas estructuras microbianas. Todos los
colorantes utilizados en microbiología tienen en común la presencia de grupos
cromóforos, grupos con dobles enlaces conjugados que son los responsables del color
mediante la unión a estructuras celulares por enlaces iónicos, covalentes o hidrófobos,
los que establecen enlaces iónicos son los más frecuentes. Entre los colorantes que se
unen por enlaces covalentes destaca el reactivo de Schiff, utilizado para la tinción de
DNA, donde el colorante se une a la desoxi-ribosa.
Por otra parte, en la tinción negativa se utilizan compuestos que no penetran en
las células sino que impregnan el medio circundante. Los microorganismos aparecen
refringentes sobre un fondo negro.
Método de la gota pendiente
Este procedimiento es la forma más sencilla de observación de organismos
vivos, nos proporciona información sobre la morfología, tamaño, color y movilidad de
las bacterias. Sin embargo, debido a la falta de contraste con el entorno es una
práctica que se utiliza únicamente para la observación del movimiento.
Tinción negativa
No se trata de una tinción propiamente dicha ya que no se utilizan colorantes
con cromóforos, ni se lleva a cabo ninguna reacción entre estructuras celulares y los
colorantes. En este caso el frotis se hace con una gota de una solución de nigrosina o
tinta china, ambos son productos particulados, insolubles, que formarán una película
opaca a la luz. En este tipo de tinción se prescinde de la fijación por calor, se deja secar
la preparación y se observan los microorganismos brillantes sobre un fondo oscuro
Tinción simple
La mayoría de los colorantes utilizados en las tinciones positivas son colorantes
derivados de las anilinas, sales orgánicas intensamente pigmentadas que proceden del
alquitrán.
Se denominan colorantes básicos si el cromóforo (porción coloreada) de la
molécula está cargada positivamente, por ejemplo, cristal violeta (Fig. 5) y azul de
metileno son colorantes básicos. Otros colorantes de esta categoría utilizados con
frecuencia en bacteriología son safranina, fuchina básica y verde malaquita. Bajo
condiciones normales de crecimiento, la mayor parte de los procariotas tienen un pH
interno próximo a la neutralidad (pH 7.0) y una superficie celular cargada
negativamente, así los colorantes básicos son los más eficaces. Colorantes ácidos con
rojo Congo, rosa de bengala, eosina y fuchina ácida tiene un cromóforo cargado
negativamente y son utilizados para teñir positivamente ciertos componentes como
las proteínas.
Tinción de Gram
Esta tinción diferencial fué propuesta por el medico danés Christian Gram (1884)
(TORTORA et al., 2007) que examinando tejido pulmonar de enfermos fallecidos por
neumonía observó que la bacteria Streptococcus pneumoniae, presente en los
pulmones, retenía el colorante conocido como marrón Bismark (sustituido
posteriormente por el cristal violeta), mientras que el tejido pulmonar no era capaz de
retener el colorante.
Es la tinción diferencial más utilizada de forma rutinaria y prácticamente la
primera prueba a la que se someten las muestras de cualquier origen antes de su
estudio. Proporciona una información esencial, además de sobre la forma, tamaño y agrupación celular, como es el tipo y composición de la pared que presentan las
bacterias. La tinción de Gram divide a las bacterias con pared del Dominio Bacteria en
dos grandes grupos: bacterias con pared de tipo gram positiva y bacterias con pared de
tipo gram negativa.
La tinción es un método sencillo para incrementar el contraste entre la célula y
su entorno y por lo tanto contribuye a mejorar la imagen observada. Las técnicas de
tinción con diversos colorantes facilitan la observación al aumentar notablemente el
contraste.
En Microbiología todas las tinciones se realizan a partir de suspensiones de
microorganismos extendidas en un portaobjetos (frotis), secadas y fijadas. La fijación,
procedimiento que permite preservar estructuras celulares, se puede llevar a cabo condiferentes tratamientos: fijación por calor o fijación química. La fijación por calor es la
más utilizada para la observación de bacterias. Este procedimiento consiste en pasar el
portaobjetos, con la suspensión bacteriana extendida y seca, a través de una llama de
un mechero. La fijación por calor preserva la morfología externa de los
microorganismos pero no las estructuras internas. La fijación química con agentes
como etanol, folmaldehido y ácido acético entre otros muchos, se utiliza para
preservar las estructuras celulares.
Se pueden utilizar dos tipos de procedimientos, la tinción positiva es la más
frecuente, en ella un colorante se une a ciertas estructuras microbianas. Todos los
colorantes utilizados en microbiología tienen en común la presencia de grupos
cromóforos, grupos con dobles enlaces conjugados que son los responsables del color
mediante la unión a estructuras celulares por enlaces iónicos, covalentes o hidrófobos,
los que establecen enlaces iónicos son los más frecuentes. Entre los colorantes que se
unen por enlaces covalentes destaca el reactivo de Schiff, utilizado para la tinción de
DNA, donde el colorante se une a la desoxi-ribosa.
Por otra parte, en la tinción negativa se utilizan compuestos que no penetran en
las células sino que impregnan el medio circundante. Los microorganismos aparecen
refringentes sobre un fondo negro.
Método de la gota pendiente
Este procedimiento es la forma más sencilla de observación de organismos
vivos, nos proporciona información sobre la morfología, tamaño, color y movilidad de
las bacterias. Sin embargo, debido a la falta de contraste con el entorno es una
práctica que se utiliza únicamente para la observación del movimiento.
Tinción negativa
No se trata de una tinción propiamente dicha ya que no se utilizan colorantes
con cromóforos, ni se lleva a cabo ninguna reacción entre estructuras celulares y los
colorantes. En este caso el frotis se hace con una gota de una solución de nigrosina o
tinta china, ambos son productos particulados, insolubles, que formarán una película
opaca a la luz. En este tipo de tinción se prescinde de la fijación por calor, se deja secar
la preparación y se observan los microorganismos brillantes sobre un fondo oscuro
Tinción simple
La mayoría de los colorantes utilizados en las tinciones positivas son colorantes
derivados de las anilinas, sales orgánicas intensamente pigmentadas que proceden del
alquitrán.
Se denominan colorantes básicos si el cromóforo (porción coloreada) de la
molécula está cargada positivamente, por ejemplo, cristal violeta (Fig. 5) y azul de
metileno son colorantes básicos. Otros colorantes de esta categoría utilizados con
frecuencia en bacteriología son safranina, fuchina básica y verde malaquita. Bajo
condiciones normales de crecimiento, la mayor parte de los procariotas tienen un pH
interno próximo a la neutralidad (pH 7.0) y una superficie celular cargada
negativamente, así los colorantes básicos son los más eficaces. Colorantes ácidos con
rojo Congo, rosa de bengala, eosina y fuchina ácida tiene un cromóforo cargado
negativamente y son utilizados para teñir positivamente ciertos componentes como
las proteínas.
Tinción de Gram
Esta tinción diferencial fué propuesta por el medico danés Christian Gram (1884)
(TORTORA et al., 2007) que examinando tejido pulmonar de enfermos fallecidos por
neumonía observó que la bacteria Streptococcus pneumoniae, presente en los
pulmones, retenía el colorante conocido como marrón Bismark (sustituido
posteriormente por el cristal violeta), mientras que el tejido pulmonar no era capaz de
retener el colorante.
Es la tinción diferencial más utilizada de forma rutinaria y prácticamente la
primera prueba a la que se someten las muestras de cualquier origen antes de su
estudio. Proporciona una información esencial, además de sobre la forma, tamaño y agrupación celular, como es el tipo y composición de la pared que presentan las
bacterias. La tinción de Gram divide a las bacterias con pared del Dominio Bacteria en
dos grandes grupos: bacterias con pared de tipo gram positiva y bacterias con pared de
tipo gram negativa.
Tinción de cápsulas
La cápsula es una estructura que rodea externamente la pared de algunos
microorganismos y constituye una barrera física que protege a la célula del ambiente
externo, proporcionando numerosas ventajas a los microorganismos que son capaces
de sintetizarla, por ejemplo, en las bacterias patógenas la cápsula permite la adhesión
al hospedador o protege de la fagocitosis, en otros casos incrementa las posibilidades
de sobrevivir en condiciones de desecación en bacterias que colonizan ambientes
naturales, o bien previenen el ataque de algunos virus, en otras ocasiones son
importantes factores de virulencia en algunas especies clínicamente relevantes como
es el caso de Streptococcus pneumoniae y también proporcionan importantes ventajas
al facilitar la adhesión a superficies sólidas. El procedimiento para la realización de la
tinción de cápsulas es el mismo que se ha explicado para la tinción negativa. Las
cápsulas aparecen rodeando a las células como estructuras sin teñir sobre un fondo
oscuro
Tinción de esporas
Esta tinción permite poner de manifiesto la endospora bacteriana, una forma de
resistencia producida por algunos géneros de bacterias gram positivas. Las endosporas
aseguran la supervivencia de estas bacterias en condiciones desfavorables (altas
temperaturas, desecación, radiaciones ultravioletas, gamma y agentes químicos),
durante largos periodos de tiempo.
METABOLISMO BACTERIANO
Las principales funciones del metabolismo son:

El metabolismo de las bacterias tiene muchos procesos en común con el metabolismo de las células eucariotas, pero algunos procesos son exclusivos del metabolismo bacteriano
Algunas particularidades del metabolimo bacteriano son:
Algunas particularidades del metabolimo bacteriano son:
El metabolismo de las bnacterias es bastante complejo. Mediante unas dos mil reacciones metabólicas diferentes la bacteria puede sintetizarse a sí misma y puede generar energía para procesos como transporte activo, motilidad y otros procesos.
Tipos de metabolismo
Los distintos tipos de metabolismo microbiano se pueden clasificar según tres criterios distintos:
1.según la forma en la que el organismo obtiene el carbono para la construcción de la masa celular:
2. según la forma en la que el organismo obtiene los equivalentes reductores para la conservación de la energía o en las reacciones biosintéticas:
3. según la forma en la que el organismo obtiene la energía para vivir y crecer:
Por lo tanto existen distintos tipos de organismos según como aprovechan el carbono y el tipo de energía que utilizan:
Los quimiolitoautótrofos obtienen energía de la oxidación de compuestos inorgánicos y el carbono de la fijación del dióxido de carbono. Ejemplos: bacterias nitrificantes, bacterias oxidantes del azufre, bacterias oxidantes del hierro, bacterias oxidantes del hidrógeno.
Los fotolitoautótrofos obtienen energía de la luz y el carbono de la fijación del dióxido de carbono, usando compuestos inorgánicos como equivalentes reductores. Ejemplos: Cyanobacteria
Los quimiolitoheterótrofos obtienen energía de la oxidación de compuestos inorgánicos, pero no pueden fijar el dióxido de carbono. Ejemplos: algunas bacterias oxidantes del hidrógeno.
Los quimioorganoheterótrofos obtienen energía, carbono y equivalentes reductores para las reacciones biosintéticas de compuestos orgánicos. Ejemplos: la mayoría de las bacterias, como Escherichia coli, Bacillus spp.,
Los fotoorganotrofos obtienen energía de la luz y el carbono y los equivalentes reductores para las reacciones biosintéticas de compuestos orgánicos.
Los quimiolitoautótrofos obtienen energía de la oxidación de compuestos inorgánicos y el carbono de la fijación del dióxido de carbono. Ejemplos: bacterias nitrificantes, bacterias oxidantes del azufre, bacterias oxidantes del hierro, bacterias oxidantes del hidrógeno.
Los fotolitoautótrofos obtienen energía de la luz y el carbono de la fijación del dióxido de carbono, usando compuestos inorgánicos como equivalentes reductores. Ejemplos: Cyanobacteria
Los quimiolitoheterótrofos obtienen energía de la oxidación de compuestos inorgánicos, pero no pueden fijar el dióxido de carbono. Ejemplos: algunas bacterias oxidantes del hidrógeno.
Los quimioorganoheterótrofos obtienen energía, carbono y equivalentes reductores para las reacciones biosintéticas de compuestos orgánicos. Ejemplos: la mayoría de las bacterias, como Escherichia coli, Bacillus spp.,
Los fotoorganotrofos obtienen energía de la luz y el carbono y los equivalentes reductores para las reacciones biosintéticas de compuestos orgánicos.

La mayoría de las bacterias patógenas son parásitos heterótrofos de seres humanos o de otras especies eucariontes. Los microorganismos heterótrofos son extremadamente abundantes en naturaleza y responsables de la degradación de los polímeros orgánicos tales como celulosa, quitina o lignina que son generalmente indigeribles para los animales más grandes. Esta degradación, generalmente, requiere la colaboración de varios organismos distintos, cada uno de los cuales realiza uno de los pasos de la degradación hasta obtener dióxido de carbono
Bioquímicamente, el metabolismo heterótrofo procariota es mucho más versátil que el de los organismos eucariotas, aunque muchos procariotas comparten los modelos metabólicos más básicos con los eucariotas, por ejemplo, usando la glicolisis para el metabolismo del azúcar y elciclo del ácido cítrico o Ciclo de Krebs en la degradacción del acetato, produciendo energía bajo la forma de ATP y reduciendo energía bajo la forma de NADH . Estas rutas metabólicas básicas están muy extendidas porque también están implicadas en la biosíntesis de muchos componentes necesarios para el crecimiento de la célula (a veces en la dirección contraria). Sin embargo, muchas bacterias utilizan rutas metabólicas alternativas
Bioquímicamente, el metabolismo heterótrofo procariota es mucho más versátil que el de los organismos eucariotas, aunque muchos procariotas comparten los modelos metabólicos más básicos con los eucariotas, por ejemplo, usando la glicolisis para el metabolismo del azúcar y elciclo del ácido cítrico o Ciclo de Krebs en la degradacción del acetato, produciendo energía bajo la forma de ATP y reduciendo energía bajo la forma de NADH . Estas rutas metabólicas básicas están muy extendidas porque también están implicadas en la biosíntesis de muchos componentes necesarios para el crecimiento de la célula (a veces en la dirección contraria). Sin embargo, muchas bacterias utilizan rutas metabólicas alternativas
Catabolismo y Anabolismo
Dado que las reacciones químicas que ocurren en la célula liberan o consumen energía, el metabolismo se puede dividir en dos clases de reacciones: catabólicas y anabólicas
Catabolismo: consiste en la degradación enzimática de macromoléculas como lípidos, hidratos de carbono y proteínas que el organismo obtiene del entorno en el que vive o de sus propias sustancias de reserva. Esta degradación se acompaña de la liberación de una gran cantidad de energía, ya que a partir de estas macromolléculas de gran complejidad estructural y alto contenido energético se obtienen moléculas sencillas estructuralmente y de bajo contenido energético. La energía que se libera durante el catabolismo es captada por la célula en forma de ATP que tiene enlaces fosfatos ricos en energía.
Anabolismo: es el proceso inverso, es la síntiesis enzimática de macromoléculas a partir de compuestos sencillos, con consumo de energía. Es decir que a partir de moléculas sencillas de escasa complejidad estructural y bajo contenido energético se sintetizan macromoléculas complejas, ricas en energía.
Las reacciones catabólicas aportan las materias primas y la energía necesaria para las reacciones anabólicas. Este acoplamiento de reacciones que liberan energía y otras que requieren energía es posible gracias al ATP (adenosintrifosfato). Las moléculas de ATP almacenan la energía proveniente de las reacciones catabólicas y la liberan en una fase ulterior para impulsar las reacciones anabólicas y el cumplimiento de otras tareas celulares. Mucha de la energía producida en el catabolismo se disipa en forma de calor.
Ambos procesos, anabólicos y catabólicos, se dan en la célula en forma simultánea e interdenpendiente.
Nutricion Bacteriana
Cualquier ser vivo, por su actividad vital (crecimiento, mantenimiento y reproducción) requiere continuos aportes de energía para reponer las pérdidas y, para que todo el sistema pueda funcionar. La nutrición es el proceso por el que los seres vivos toman del medio donde habitan las sustancias químicas que necesitan para crecer. Dichas sustancias se denominan nutrientes, y se requieren para los dos objetivos que comprende el metabolismo:
El estudio de la nutrición microbiana se puede desglosar en varios apartados: así, podemos considerar los tipos de nutrientes requeridos, los aspectos cuantitativos, e incluso podemos abordar los aspectos ambientales (en cuyo caso entramos dentro del campo de la Ecofisiología). Igualmente podemos estudiar la aplicación práctica de la nutrición bacteriana, que se plasma sobre todo en el diseño de medios de cultivo para manejar los microorganismos en el laboratorio.
Es importante tener claros desde el principio una serie de conceptos y nomenclaturas relacionados con los principales tipos de nutrición bacteriana. Puesto que, como acabamos de ver, la nutrición presenta un aspecto de aprovisionamiento de energía y otro de suministro de materiales para la síntesis celular, podemos hablar de dos "clasificaciones" de tipos de nutrición:
Desde el punto de vista de los fines de aprovisionamiento de energía, las bacterias se pueden dividir en:
Sean autótrofas o heterótrofas, todas las bacterias necesitan captar una serie de elementos químicos, que se pueden clasificar (según las cantidades en que son requeridos) como
En la naturaleza, estos elementos se encuentran combinados, formando parte de sustancias orgánicas y/o inorgánicas. Algunos de los nutrientes serán incorporados para construir macromoléculas y estructuras celulares; otros solo sirven para la producción de energía, y no se incorporan directamente como material celular; finalmente, otros pueden ejercer ambos papeles.
El mundo bacteriano, como conjunto, exhibe una gigantesca versatilidad metabólica de uso de nutrientes: desde autótrofos que obtienen su carbono por reducción del CO2 y los demás elementos a partir de fuentes igualmente inorgánicas, hasta heterótrofos capaces de usar amplia gama de fuentes orgánicas de carbono.
A su vez, dentro de los heterótrofos, podemos encontrar muchos tipos de nutrición muy distintos, desde bacterias metilotrofas que sólo usan metano o metanol como fuente de carbono y energía, hasta los muy versátiles Pseudomonas, que pueden recurrir a degradar más de 100 tipos de fuentes de C, incluyendo sustancias tan "exóticas" como hidrocarburos alifáticos y cíclicos. De cualquier modo, entre los heterótrofos, una de las fuentes más típicas de carbono consiste en glucosa.
CLASES DE NUTRIENTES
Podemos clasificar los nutrientes en las siguientes categorías:
NUTRIENTES PARTICULARES
Se trata de elementos que pueden ser cubiertos de modo muy distinto, dependiendo del tipo de bacteria que consideremos. Concretamente, los elementos N y S (que requieren todos los seres vivos) pueden ser captados por las bacterias de modos muy distintos, dependiendo de sus capacidades biosintéticas.
Cualquier ser vivo, por su actividad vital (crecimiento, mantenimiento y reproducción) requiere continuos aportes de energía para reponer las pérdidas y, para que todo el sistema pueda funcionar. La nutrición es el proceso por el que los seres vivos toman del medio donde habitan las sustancias químicas que necesitan para crecer. Dichas sustancias se denominan nutrientes, y se requieren para los dos objetivos que comprende el metabolismo:
- fines energéticos o catabolismo (reacciones de mantenimiento)
- fines biosintéticos o anabolismo (reacciones plásticas ).
El estudio de la nutrición microbiana se puede desglosar en varios apartados: así, podemos considerar los tipos de nutrientes requeridos, los aspectos cuantitativos, e incluso podemos abordar los aspectos ambientales (en cuyo caso entramos dentro del campo de la Ecofisiología). Igualmente podemos estudiar la aplicación práctica de la nutrición bacteriana, que se plasma sobre todo en el diseño de medios de cultivo para manejar los microorganismos en el laboratorio.
Es importante tener claros desde el principio una serie de conceptos y nomenclaturas relacionados con los principales tipos de nutrición bacteriana. Puesto que, como acabamos de ver, la nutrición presenta un aspecto de aprovisionamiento de energía y otro de suministro de materiales para la síntesis celular, podemos hablar de dos "clasificaciones" de tipos de nutrición:
Desde el punto de vista de los fines de aprovisionamiento de energía, las bacterias se pueden dividir en:
- litotrofas (del griego lithos = piedra): son aquellas que sólo requieren sustancias inorgánicas sencillas (SH2 , S0, NH3, NO2-, Fe, etc.).
- organotrofas: requieren compuestos orgánicos (hidratos de carbono, hidrocarburos, lípidos, proteínas, ......).
- autótrofas: crecen sintetizando sus materiales a partir de sustancias inorgánicas sencillas. Ahora bien, habitualmente el concepto de autotrofía se limita a la capacidad de utilizar una fuente inorgánica de carbono, a saber, el CO2.
- heterótrofas: su fuente de carbono es orgánica (si bien otros elementos distintos del C pueden ser captados en forma inorgánica).
| autótrofas estrictas: son aquellas bacterias incapaces de crecer usando materia orgánica como fuente de carbono. | |
| mixótrofas son aquellas bacterias con metabolismo energético litótrofo, pero requieren sustancias orgánicas como nutrientes para su metabolismo biosintético. |
| macronutrientes (C, H, O, N, P, S, K, Mg) | |
| micronutrientes o elementos traza (Co, Cu, Zn, Mo...) |
En la naturaleza, estos elementos se encuentran combinados, formando parte de sustancias orgánicas y/o inorgánicas. Algunos de los nutrientes serán incorporados para construir macromoléculas y estructuras celulares; otros solo sirven para la producción de energía, y no se incorporan directamente como material celular; finalmente, otros pueden ejercer ambos papeles.
El mundo bacteriano, como conjunto, exhibe una gigantesca versatilidad metabólica de uso de nutrientes: desde autótrofos que obtienen su carbono por reducción del CO2 y los demás elementos a partir de fuentes igualmente inorgánicas, hasta heterótrofos capaces de usar amplia gama de fuentes orgánicas de carbono.
A su vez, dentro de los heterótrofos, podemos encontrar muchos tipos de nutrición muy distintos, desde bacterias metilotrofas que sólo usan metano o metanol como fuente de carbono y energía, hasta los muy versátiles Pseudomonas, que pueden recurrir a degradar más de 100 tipos de fuentes de C, incluyendo sustancias tan "exóticas" como hidrocarburos alifáticos y cíclicos. De cualquier modo, entre los heterótrofos, una de las fuentes más típicas de carbono consiste en glucosa.
En los heterótrofos- organotrofos, los sustratos carbonados (con un nivel de oxidación no muy distinto del material celular -CH2O-) entran simultáneamente al:
| metabolismo energético (o catabolismo, donde la fuente de C se transforma en CO2, o en CO2 junto con otras sustancias no totalmente oxidadas); | |
| metabolismo plástico (anabolismo = biosíntesis de nuevo material celular). |
Aunque dentro del mundo de los procariotas se encuentre tanta variedad de nutriciones, las bacterias que pueden nutrirse solamente de sustancias inorgánicas sencillas (H2O, CO2, N2, NO3--, NH3, SO4=, fosfatos, etc.) son minoría, pero sus procesos metabólicos son muy interesantes. De hecho, existen tipos metabólicos que sólo han evolucionado en procariotas.
Como paradigma de esto citaremos los microorganismos quimioautótrofos (o quimiolitoautótrofos): obtienen su energía de la oxidación de sustancias inorgánicas sencillas, el carbono procede del CO2, y el resto de elementos a partir de sales inorgánicas, por lo que pueden vivir en soluciones de sales minerales.
Lo habitual, sin embargo, es que muchas bacterias recurran, siempre que puedan, a tomar del medio ciertos compuestos más complejos, ya que carecen de ciertas rutas biosintéticas.
CLASES DE NUTRIENTES
Podemos clasificar los nutrientes en las siguientes categorías:- universales: agua, CO2, fosfatos y sales minerales;
- particulares
- factores de crecimiento
NUTRIENTES PARTICULARES
Se trata de elementos que pueden ser cubiertos de modo muy distinto, dependiendo del tipo de bacteria que consideremos. Concretamente, los elementos N y S (que requieren todos los seres vivos) pueden ser captados por las bacterias de modos muy distintos, dependiendo de sus capacidades biosintéticas.
Tanto el N como el S se encuentran en la célula en estado reducido:
| el radical -NH2 forma parte de los aminoácidos (que a su vez son los sillares de las proteínas) y de las bases nitrogenadas (que participan en los ácidos nucleicos y en algunas coenzimas); | |
| el radical -SH interviene en determinados aminoácidos y en coenzimas como la CoA. |
¿En qué formas químicas entran N y S a las bacterias?
La mayoría de bacterias fotosintéticas y muchas heterotrofas asimilan estos elementos en forma combinada inorgánica oxidada:
| como NO3--, merced a la actuación secuencial de nitrato- reductasas y nitrito- reductasas asimilatorias | |
| como SO4=. Este sulfato se activa con ATP, y luego se reduce hasta sulfito y finalmente sulfhídrico, que ya tiene el estado de reducción adecuado para la incorporación del S. |
Muchas bacterias heterótrofas pueden usar alguna forma reducida:
| de N inorgánico: amonio (NH4+) | |
| de S inorgánico: sulfuros (S=, SH-) | |
| de N orgánico: aminoácidos, péptidos | |
| de S orgánico: cisteína. |
Muchas de las bacterias que pueden usar amonio como única fuente de nitrógeno también pueden usar nitratos.
Nutrición bacterianaBuscar en el sitio:CONCEPTOS BÁSICOS | CLASES DE NUTRIENTES | NUTRIENTES UNIVERSALES | NUTRIENTES PARTICULARES | FIJACIÓN DE NITRÓGENO | FACTORES DE CRECIMIENTO | MEDIOS DE CULTIVO |GLOSARIO | BIBLIOGRAFÍA | AUTO- EVALUACIÓN
CONCEPTOS BÁSICOS
Cualquier ser vivo, por su actividad vital (crecimiento, mantenimiento y reproducción) requiere continuos aportes de energía para reponer las pérdidas y, para que todo el sistema pueda funcionar. La nutrición es el proceso por el que los seres vivos toman del medio donde habitan las sustancias químicas que necesitan para crecer. Dichas sustancias se denominan nutrientes, y se requieren para los dos objetivos que comprende el metabolismo:
Es importante tener claros desde el principio una serie de conceptos y nomenclaturas relacionados con los principales tipos de nutrición bacteriana. Puesto que, como acabamos de ver, la nutrición presenta un aspecto de aprovisionamiento de energía y otro de suministro de materiales para la síntesis celular, podemos hablar de dos "clasificaciones" de tipos de nutrición: Desde el punto de vista de los fines de aprovisionamiento de energía, las bacterias se pueden dividir en:
En la naturaleza, estos elementos se encuentran combinados, formando parte de sustancias orgánicas y/o inorgánicas. Algunos de los nutrientes serán incorporados para construir macromoléculas y estructuras celulares; otros solo sirven para la producción de energía, y no se incorporan directamente como material celular; finalmente, otros pueden ejercer ambos papeles.El mundo bacteriano, como conjunto, exhibe una gigantesca versatilidad metabólica de uso de nutrientes: desde autótrofos que obtienen su carbono por reducción del CO2 y los demás elementos a partir de fuentes igualmente inorgánicas, hasta heterótrofos capaces de usar amplia gama de fuentes orgánicas de carbono. A su vez, dentro de los heterótrofos, podemos encontrar muchos tipos de nutrición muy distintos, desde bacterias metilotrofas que sólo usan metano o metanol como fuente de carbono y energía, hasta los muy versátiles Pseudomonas, que pueden recurrir a degradar más de 100 tipos de fuentes de C, incluyendo sustancias tan "exóticas" como hidrocarburos alifáticos y cíclicos. De cualquier modo, entre los heterótrofos, una de las fuentes más típicas de carbono consiste en glucosa.En los heterótrofos- organotrofos, los sustratos carbonados (con un nivel de oxidación no muy distinto del material celular -CH2O-) entran simultáneamente al:
Como paradigma de esto citaremos los microorganismos quimioautótrofos (o quimiolitoautótrofos): obtienen su energía de la oxidación de sustancias inorgánicas sencillas, el carbono procede del CO2, y el resto de elementos a partir de sales inorgánicas, por lo que pueden vivir en soluciones de sales minerales. Lo habitual, sin embargo, es que muchas bacterias recurran, siempre que puedan, a tomar del medio ciertos compuestos más complejos, ya que carecen de ciertas rutas biosintéticas.
CLASES DE NUTRIENTES
Podemos clasificar los nutrientes en las siguientes categorías:
NUTRIENTES UNIVERSALES
EL AGUALas bacterias necesitan grandes cantidades de agua. De hecho, salvo excepciones, se pueden considerar como organismos acuáticos. Requieren cierto grado de humedad para crecer. Desde el punto de vista de sus posibles papeles, el agua es:
aW= PS/PW donde PS es la presión parcial de vapor de agua en la solución problema y PW es la presión parcial de vapor del agua destilada. ¿Cómo medir el potencial de agua de un medio líquido determinado? Se mide la humedad relativa del aire que hay encima del medio (en frascos), y este valor se refiere al valor de 100, que es la humedad del aire encima del agua destilada. O sea, la aW = humedad relativa/100). Las bacterias tienen valores de aW normalmente entre 0.90 y 0.99.
EL ANHÍDRIDO CARBÓNICO (CO2)
El anhídrido carbónico es requerido por todo tipo de bacterias:
FÓSFORO
Suele requerirse en forma de fosfatos, sea orgánicos o inorgánicos. Las bacterias que pueden usar los fosfatos orgánicos (merced a la posesión de fosfatasas) no dependen absolutamente de ellos, ya que pueden recurrir igualmente a los fosfatos inorgánicos. Los fosfatos orgánicos son hidrolizados por fosfatasas extracelulares o (en las Gram-negativas) periplásmicas (p.ej., la fosfatasa alcalina).El fósforo se usa principalmente para la síntesis de los ácidos nucleicos y los fosfolípidos, pero aparece también encoenzimas y en proteínas.
SALES MINERALES
Las sales minerales son la fuente de aniones (p. ej. el Cl-) y de cationes para la célula. Los siguientes cationes, concretamente, se necesitan en cantidades relativamente grandes: K+, Mg++, Ca++, Fe++.El ión potasio (K+):
El hierro (principalmente como ión ferroso, Fe++) suele estar acomplejado en la naturaleza, formando sales insolubles. Las bacterias disponen de una serie de moléculas, denominadas sideróforos, capaces de captar ese hierro. El hierro:
NUTRIENTES PARTICULARES
Se trata de elementos que pueden ser cubiertos de modo muy distinto, dependiendo del tipo de bacteria que consideremos. Concretamente, los elementos N y S (que requieren todos los seres vivos) pueden ser captados por las bacterias de modos muy distintos, dependiendo de sus capacidades biosintéticas.Tanto el N como el S se encuentran en la célula en estado reducido:
La mayoría de bacterias fotosintéticas y muchas heterotrofas asimilan estos elementos en forma combinada inorgánica oxidada:
FIJACIÓN DE NITRÓGENO
La atmósfera contiene enormes cantidades de nitrógeno no combinado (libre) en estado gaseoso: el nitrógeno molecular o dinitrógeno (N2), que procede de microorganismos denitrificantes (véase el capítulo 15). Sin embargo, esta gran reserva sólo puede servir de fuente de nitrógeno a ciertos procariotas, las llamadas bacterias fijadoras de nitrógeno o diazotrofos. Esta notable capacidad bioquímica no ha evolucionado en eucariotas. (Lo más a que ha llegado la evolución es a seleccionar ciertos tipos de asociaciones simbióticas entre procariotas diazotrofos y ciertos eucariotas). En una época todo el nitrógeno de los seres vivos fue procesado por estas bacterias, que toman el nitrógeno atmosférico (N2) y lo modifican en manera tal que pueden ser utilizados por los organismos vivos como nitratos o amoníaco NH3.Desarrollo de un nódulo en una raíz de ciertas plantas (en general leguminosas) donde viven bacterias en simbiosis con ella    La fijación del N2, es un proceso de reducción que convierte el nitrógeno molecular en amoniaco, según la siguiente ecuación: N2 + 8H+ + 8e + 18 ATP -->2NH3 + H2 + 18 (ADP + Pi)Esta reacción está catalizada por un complejo enzimático denominado nitrogenasa o dinitrogenasa, que consta de dos componentes:
Un aspecto muy importante del complejo nitrogenasa es su extrema sensibilidad al oxígeno, de modo que queda rápida e irreversiblemente inactivado por este gas. Ahora bien, esto no significa que la capacidad de diazotrofía esté relegada a bacterias anaerobias, ya que, la evolución ha "inventado" distintas estrategias para proteger a la nitrogenasa en fijadores aerobios como las cianobacterias que han confinado el sistema al interior de células con gruesas paredes (los heterocistos). Debido a lo "caro" que resulta fijar nitrógeno, no es extraño comprobar que este proceso esté regulado de forma muy estricta ante la presencia en el medio de fuentes combinadas de nitrógeno (nitratos, amonio, aminoácidos):
FACTORES DE CRECIMIENTO
Los factores de crecimiento son moléculas orgánicas específicas que, en muy pequeña cantidad, algunas bacterias necesitan para crecer. Salvo excepciones no tienen función plástica (no son sillares de macromoléculas) ni sirven como fuente de energía. Suelen ser coenzimas o sus precursores, vitaminas, que determinadas bacterias no pueden fabricar por sí mismas, al carecer de parte o toda de una ruta biosintética.Ejemplos:
MEDIOS DE CULTIVO Y CONDICIONES DE CRECIMIENTO
El conocimiento de la nutrición microbiana permite el cultivo de los microorganismos en el laboratorio. Como acabamos de ver, en general, todos los microorganismos tienen parecidos requerimientos de macro- y micronutrientes, aunque ha quedado claro que la forma en que cada nutriente es captado puede variar mucho entre unas bacterias y otras, así como la cantidad relativa de cada nutriente. Los microbiólogos, en su trabajo cotidiano, están acostumbrados a manejar multitud de "recetas" o fórmulas correspondientes a muchos tipos de medios de cultivo.Un medio de cultivo es una solución acuosa (bien como tal, o bien incorporada a un coloide en estado de gel) en la que están presentes todas las sustancias necesarias para el crecimiento de un (os) determinado (s) microorganismo (s). Los medios de cultivo se pueden clasificar, en primera instancia, en tres grandes tipos: 1) Medios complejos o indefinidos: su composición química exacta se desconoce, ya que son el producto de realizar infusiones y extractos de materiales naturales complejos:Ejemplos:
2) Medios sintéticos o definidos: se obtienen disolviendo en agua destilada cantidades concretas de distintas sustancias químicas puras, orgánicas y/o inorgánicas. La composición concreta de un medio sintético dependerá de la bacteria que queramos cultivar: lógicamente, un medio definido para una bacteria con grandes capacidades biosintéticas será más sencillo que el medio definido de otra bacteria con menores posibilidades biosintéticas. 3) En tercer lugar, podemos fabricar un tipo de medio "mezcla" de los anteriores, denominado medio semisintético: llevan algunas sustancias químicas cuya naturaleza y cantidad conocemos, junto con sustancias de naturaleza y composición indefinidas. Cualquiera que sea el tipo de medio, de los tres que acabamos de citar, podemos elaborar dos tipos de "versiones", según su estado aparente:
|
Suscribirse a:
Entradas (Atom)